|
分 類: |
|
|
|
1. |
Pythium spp.
Oomycetes (卵菌綱)
Peronosporales (露菌目)
Pythiaceae (腐霉菌科)
Pythium (腐霉菌屬) |
|
|
2. |
Fusarium oxysporum f.sp. lilii
Deuteromycetes (不完全菌綱)
Moniliales (線菌目)
Tuberculariaceae (瘤座孢科)
Fusarium (鐮孢菌屬) |
|
|
3. |
Rhizoctonia solani
無性世代
Deuteromycetes (不完全菌綱)
Agonomycetales (無孢子菌目)
Mycelial sterilia (無孢子菌科)
Rhizoctonia (絲核菌屬)
有性世代
Basidiomycetes (擔子菌綱)
Tulasnellales (膠膜菌目)
Ceratobasidiaceae (角擔菌科)
Thanatephorus |
|
分 佈: |
全世界百合種植區均可發現,但主要分佈在較溫暖的地帶,在美國本病為百合生長之限制因子。本病主要是由多種土壤傳播性真菌複合感染所致,但不同地區主要發生之病原真菌並不相同。一般而言,排水不良地區或水份潛勢
(water potential) 較低之地方以 Pythium spp. 及 R. solani 發生較多,而
F. oxysporum 則在任何土壤濕度下均可危害。 |
|
|
1. |
Pythium spp. :
根據報告自罹病百合根部曾經分離出之腐霉菌 ( Pythium spp.) 包括 P. spinosum、P.
ultimum 及 P. splendens。在歐洲地區、美國加州、明尼蘇達及奧勒岡州腐霉菌之發生以
P. ultimum 為主。P. splendens 則發生在美國佛羅里達州,臺灣所發生者則為P.spinosum。 |
|
|
2. |
Fusarium oxysporum:
本病原菌係一種相當普遍之土棲性真菌,存在於溫帶及熱帶之耕作土壤中,包括腐生性及具病原性者。本菌可經由種球帶菌或直接由土壤侵害百合,尤其在高溫多濕之季節發生最為嚴重。 |
|
|
3. |
Rhizoctonia solani
R. solani分佈世界各地,並可危害多種作物,任何適合耕作之土壤均可能含有本菌。本菌屬可依其菌絲融合之特性區分成不同之菌絲融合群
(Anastomosis group,簡稱 AG)。在臺灣罹病百合根部上所發現者為 AG-1 , AG-3 及 AG-4
等三群,而以 AG-4 群為害最鉅。 |
|
寄 主: |
F. oxysporum、R.
solani 及 Pythium spp. 均為多犯性植物病原真菌,且為重要的非專一性土壤傳播性病原菌。Pythium spp. 寄主包括茄科、葫蘆科、薑科、豆科、禾本科及花卉等。R. solani
可侵害 66 科約 300
種植物。AG-4
群之寄主包括甜菜、柑桔、棉花、苜蓿、菜豆、大豆、松、落花生、十字花科、蔬菜、洋蔥、菊、茄科作物、亞麻、鐘麻及西瓜等。
F. oxysporum 有 75 種以上之分化型,每個分化型都有獨特之寄主範圍,引起百合根腐者為 F. oxysporum f. sp.
lilii。 |
|
形 態: |
|
|
|
1. |
Pythium spp. :
菌絲無色、透明、無隔膜,成不規則之樹枝狀。孢囊 (sporangium)
之大小及形狀在種間有很大差異,可作為分類之依據。P. ultimum及P. splendens
為分生孢子型,P.
spinosum 則為圓球型。此三種之孢囊為頂生或中生。藏卵器為球型,主要形成於菌絲頂端,P. spinosum
者則為中生型,且器壁有點狀突起,其餘二種則表面平滑,每個藏卵器有 1-6 個藏精器著生於下方。P. ultimum及P.
spinosum只有 1-2 個,P. splendens 則有 2-4 個。 |
|
|
2. |
Fusarium oxysporum :
大孢子 (macroconidium),3-5個隔膜,薄壁,彎曲幅度不大,有足型之底細胞 (basal
cell),小孢子 (microconidium) 透明,通常為單室,卵型至腎形,只以假頭狀產生於短孢子柄上。厚膜孢子
(chlamydospores) 單一或成雙形成。在馬鈴薯葡萄糖培養基上之菌落呈淡紫色。 |
|
|
3. |
Rhizoctonia solani :
R. solani 具備五項形態特徵:營養菌絲細胞多核;菌絲隔板具有隔膜孔 (dolipore septum);
菌絲分支經常發生於主軸菌絲前端之隔板附近; 分支菌絲基部會縊縮,距主軸菌絲不遠處有隔板; 菌叢呈褐色等。R. solani
之無性世代不產生無性孢子,僅以菌絲和菌核繁殖。菌絲細胞核之數目為區別 Rhizoctonia 屬之依據,R. solani
菌絲為多核型。菌絲培養時生長快速,初期為白色,而後轉變呈褐色,菌絲寬度為
6-12μm,分枝菌絲常與原菌絲呈很大角度
(約呈直角)。菌核暗色不規則型,可在被害植物組織中大量產生,由念珠狀細胞構成,內部常有營養菌絲存在,大小依培養條件及不同菌株而有差異。菌核為越冬的菌體,直接發芽後形成菌絲。 |
|
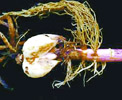
診斷技術: |
本病害主要發生於百合根部,在發病初期植株並無明顯病徵,當植株顯現下位葉黃化萎凋病徵時,已為感染末期,此時根系大多數已腐爛。百合根腐病主要由三種病原真菌複合感染。因此很難由根系受害外觀判斷是由何種
(或幾種) 真菌危害所造成。粗略之判定法是若維管束已變色,則表示有 F. oxysporum
參與危害;若根系呈水浸狀,無變色則為 Pythium spp. 所引起;根上若有褐色斑點,則有可能為 R. solani
所造成。但目前唯一正確之診斷方法是將罹病根部取回,以組織分離法行表面消毒,水洗切段後置於水瓊脂培養基上,待菌絲長出後挑取尖端部位,移殖至馬鈴薯葡萄糖培養基上,培養幾天後可依菌落形態及孢子特徵來判別病原菌種類。 |
|
生 活
史: |
|
|
|
1. |
Pythium spp. :
卵孢子是 Pythium spp.
在土壤中主要之殘存構造。卵孢子發芽後所形成之菌絲可直接侵入百合根部,或是形成孢囊,再釋出游走子。游走子可借水游向根系,並在根上靜止形成小孢囊
(Encyst)
後侵入。待菌絲大量纏化罹病組織後,再形成孢囊,遇水即大量釋放游走子,進行第二次感染。藏精器及藏卵器形成於壞死組織上,經有性生殖後形成越冬之卵孢子,卵孢子發芽後再引起下一次之危害。 |
|
|
2. |
Fusarium oxysporum:
本菌可腐生於土壤中,或於植株殘體上繁殖。環境不良時可大量形成厚膜孢子作為殘存構造。菌絲侵入百合根部後,沿內、外皮層間向前蔓延,菌絲很少侵入內皮層組織內(endodermis)。除根系外,此菌亦可危害鱗片、根盤及球莖,感染後期在寄主組織表面上形成大量分生孢子,並進行第二次感染。厚膜孢子形成於環境不良時,並為殘存或越冬之工具,當遇到適宜寄主即可發芽,再侵入危害。 |
|
|
3. |
Rhizoctonia solani :
R. solani 以菌核或存活於植株殘體上之菌絲為主要感染源。菌核可產生並存活於寄主組織中,無寄主時菌核亦可存活於土壤中,寄主根部分泌物或土中有機質均可刺激其發芽。若土中有機質豐富,則本菌可行腐生生活,而長時間殘存於土中。當遇及適當寄主時,R.
solani 可產生纖維、果膠分解酵素及植物毒質 (phytotoxin)
來殺死寄主細胞,分解其組織,並促使其釋出有機養分供菌體繼續蔓延生長。一般而言,本菌在乾土中之壽命比在潮濕土壤中更長。 |