

 19767
19767
蔬菜種子活力與種子處理之探討
種苗改良繁殖場 黃玉梅
一、前言
種子活力是影響種子品質的要因,其除了以傳統的發芽率來表示其活力(Viability)外,必須輔以種子活勢(Vigour),才能反映一批種子的成株率,或貯藏能力。高活力之種子乃是種子生產者與種苗業者共同追求的目標,100%的發芽率更是他們共同的夢想。而種子活力受本身(成熟度、種子大小重量及顏色、機械傷害)與環境(溫度、日照、營養之供給影響,但不管任何具有高活力的種子,經過一段時間貯存,種子供然會逐漸老化、衰敗(細胞膜逐漸崩解、粒線體活力降低,ATP含量減少、呼吸作用逐漸降低、生他合成能力降低等現象)最後失去活力。
種子生理學者與種子實踐工作者為探求活力,而研究出各種種子處理技術:添加殺菌劑、養分、荷爾蒙及植物生長調節劑:種子造粒(披衣)(Seed pelleting)處理、粉衣和膜衣處理(Coating and film coating)、滲調處理(Priming)、流種播種(Fluid drilling)、物理性揀別等,提高種子發芽率。Taylor等學者更結合了Priming及coating技術、利用sinapine之螢光反應,直接揀除無活力之種子,有效地提高甘藍等種子發芽率,成為種子研究工作者另一個值得探討的項目。
種子是作物生產的根基,尤其是經由種子生產的作物(如蔬菜作物),種子發芽與幼苗成活之好壞直接影響作物的產量與品質。活力高的種子,發芽迅速、整齊、成活率高、幼苗對不良環境之抵抗力強,且成熟期早、產量亦高(俞,1990)。但不管在任何有利的貯藏環境下,經過一段時間,種子仍然會逐漸老化、衰敗、最後常去活力。根據統計美國每年由於種子活力下降而造成的損失在十億美元以上。然而種子老化又是不可逆的生理現象,因此,研究種子活力下降的生理、生化現象,探求提高活力的可能途徑,就成為種子生理學研究及種子實踐工作中重要的研究內容(劉,1988)。
近年來,自動化穴盤育苗生產蓬勃發展,為增加項高投資集約栽培育苗系統的成效性,減少育苗時種子需要較長發芽時間,及發芽不理想造成缺株,而耗損生產體系設施下時間、空間與人力成本。因此,使種子具有高品質與活力的調製技術或直接揀除不具活力之種子,提高種子發芽率與發芽勢成為自重化育苗工作中非常重要的一環。
種子活力的定義:早期對種子活力定義不甚明確,往往將種子活勢(Vigour)與種子存活力(Viability)視為相同,且認為種子活力等於種子發芽力。直至1950年國際種子會議才將種子活勢與存活力清楚的區分,並歸為影響種子品質的項目之一(劉,1988)。有關seed vigour 定義極多,Isiey對種子活勢認為在不良的田間環境下,能迅速發芽且整齊的能力(Woodtock, 1967)。Perry 則認為種子活勢為種子在土壤中產生幼苗的能力,以及種子適應環境因子範圍的水平,受遺傳基因決定,而能為環境所改變。
直至1977 年國際種子檢查協會(ISTA),將種子活勢的定義認種子發芽或幼苗出土(Seedling emergence)期間決定種子的表現及活性潛勢的特質(1989),包括四項特質:1.發芽期間生化過程與反應,如酵素反應與呼吸活性。2.種子發芽和幼苗生長的速度與整齊度。3.種子於田間萌發的速度與整齊度。4.在不適合的環境下,幼苗出土的能力。
至 80 年代對種子活力的研究更深入細胞胞器,酵素與核酸代謝的研究領域中,種子活力應從下列考慮幾點:1.種子吸脹水後,原生質和細胞器膜,系統修復與重組的速度與能力。2.基質與貯藏物質的降解,提供細胞組成的效率。3.蛋白質與 RNA 合成速率。4.粒腺體的合成與功能。5.蛋白質與酵素的穩定性。6.貯存質體(Plastid)的完整性。
1992年國際種子會議討論 21 篇報告後,認為種子潛力(Potential)應包含種子本身的存活率(viability)及它能適應田間環境的能力,所表現出來的活勢(Vigour),這樣才能表現一批種子的成株率。因此認為種子潛力應包括三方面即種子存活力、種子活勢、及田間能力(Field Performance),將田間能力與存活力及活勢區分成為另一項考慮因素(Miller 1994)。
二、影響種子活力的因子:
(一)環境因子:
1.溫度:
種子形成時之溫度會影響到收穫後之發芽能力,一般日、夜溫差大有助於養分蓄積,使種子充實,例如萵苣種子成熟期的高溫可明顯提高發芽能力(杉山直儀,1984),低溫對胚發育不利甚至受害。
2.日照:
種子形成時之日照長短與溫度一樣,對種子活力有很大的影響,萵苣種子形成時每天照光8小時所得之種子,其發芽能力較每天照光20小時者為高(高、湯,1977)。
3.營養之供給:
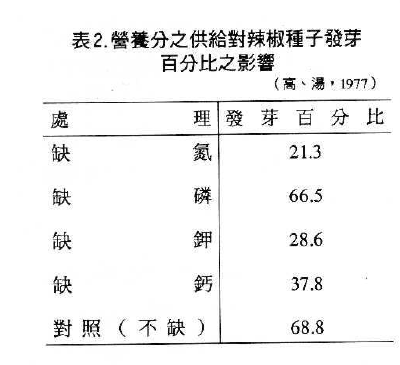
母株生育時營養分是否正常供應,不但影響到產量,且影響到採收後之發育能力。Harrington以辣椒為材料,研究嚴重缺氮、缺磷、缺鉀、缺鈣對收穫種子發芽之影響。由表2可知,除缺磷外,其餘均會降低發芽率(高與湯,1977)。
(二)種子本身:
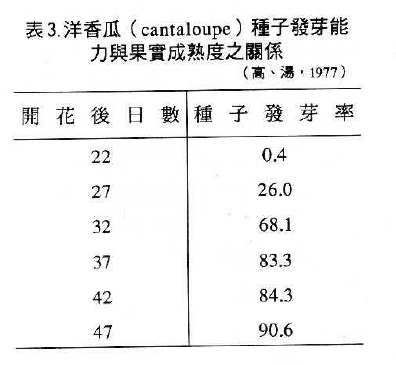
1.成熟度:
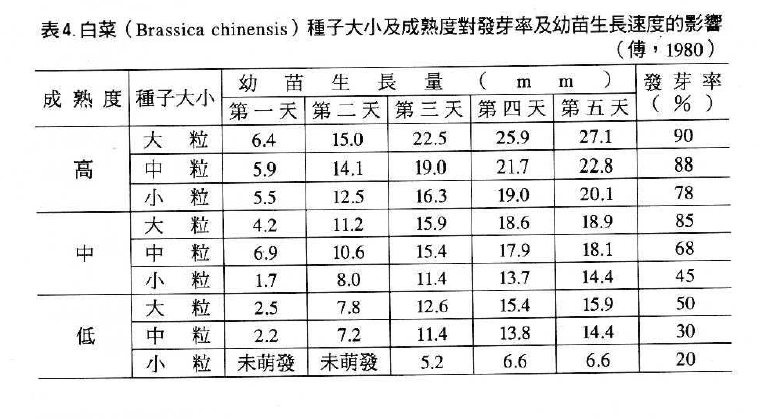
來自成熟果實之種子,其發芽力高於尚未成熟之果實如表3洋香瓜。同樣的,成熟度高的種子及種子大小均影響種子活力,如表4的白菜種子(劉,1988)。
2.種子大小重量及顏色:
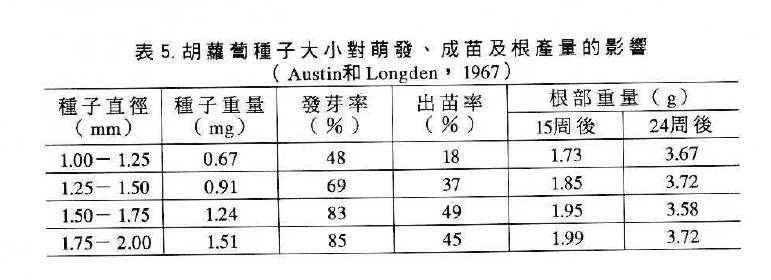
許多報告指出番茄與辣椒種子之發芽及胡蘿蔔種子在田間之萌芽與種子大小呈正相關,蘿蔔、萵苜、青花菜及胡瓜等種子大小、重量也與幼苗大小及產量呈正相關(如表5 )。
3.機械傷害:
蟲害及採收處狂的機械傷害,使種子增加病源的感染,且對生長組織(胚根、胚莖、上胚軸)及貯藏組織(胚乳)直接產生傷害,使種子活力迅速下降。
三、種子的老化與衰敗:
種子為一具有生命的有機體,因此和地球上的任何生物一樣,會隨著不可逆的老化(ageing)使得種子活力下降,終至死亡這種逐漸老化的現象我們稱之為衰敗(deterioration)。
(一)種子衰敗的生理現象:
1.細胞膜逐漸崩解:
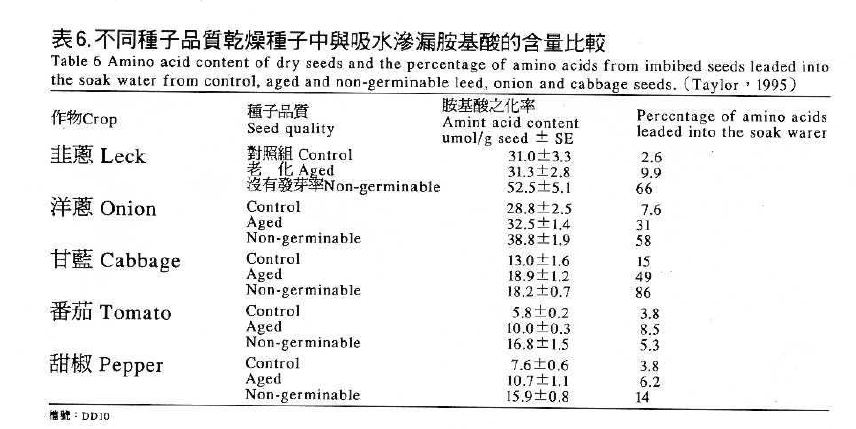
在種子老化的過程中構成細胞膜的脂質因過氧化反應產生自由基而破壞膜系,一旦細胞膜崩解後,通透性增加,使細胞內部許多大分子如醣類、氨基酸等無法被細胞膜保存於細胞內而向外滲漏(leakage),致使土壤中之微生物在種子表面獲取足夠之養分而滋生,最後使種子腐爛或幼苗死亡(俞,1981)。由表6可看出韭蔥、洋蔥、 甘藍、番茄、甜椒等五種蔬菜其老化種子,氨基酸滲漏量均比未老化之種子多,且不能發芽的種子滲漏量更多。(Taylor,1995)。
2.粒線體活力降低,ATP含量減少:
因粒線體雙層膜構造遭破袁而使粒腺體活力下降,一旦粒腺體活力下降後,使得在粒腺體內所進行的TCA cycle、電子傳遞系統、氧化磷酸化作用受阻,而使ATP含量減少。
3.吸呼作用逐漸降低:
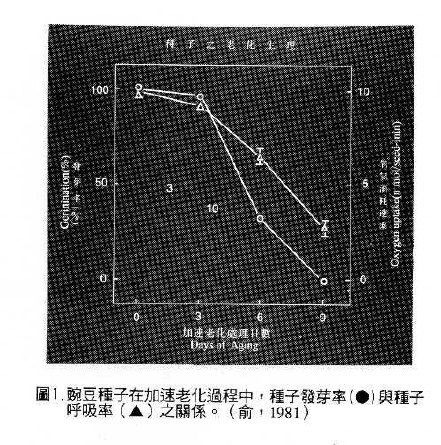
如前所述,因粒線體之活力與正常呼吸作用有極密切之關係,粒線體活力下降,使呼吸率降低。如圖1可看出新鮮種子之發芽率高,幼苗生長迅速,呼吸率亦高;發芽率低的種子,幼苗生長速率慢,其呼吸率亦隨著降低。呼吸率隆低可能限制了許多發芽時必須能量的供應(俞,1981 )。
4 .酵素活性改變:
種子老化的過程中具有破壞性的水解酵素活性增加如:蛋白酵素、磷酸化酵素、肌醇六磷酸化酵素而呼吸系統酵素活性下降如:過氧化氫酵素、過氧化物酵素、脫氫酵素、谷氨酸脫酸酵素、細胞色素氧化酵素。其中以脫氫酵素活性下降最明顯,且與種子發芽率下降速度相平行,因此常被用來檢定種子活性。
5.蛋白質能力降低:
Adbul-Baki以32P及14C-leucine加入不同老化程度之大豆種子胚中,追蹤RNA與蛋白質之合成,發現種子老化時,種子內核酸含量不因老化而改變,而是由於種子老化使DNA遭分解成小分子,而使其模板能力故障,同時進行錯誤的轉譯作用,使產生多量的小分子RNA,而抑制蛋白質之合成(王,1986)。另外,因大分子的合成為一種需能反應,種子老化時亦可使蛋白質合成能力降低。
(二)種子老化之生理機制:
引起種子老化的重大原因乃細胞膜以及細胞內各質體膜之變化。細胞膜之組成分不外乎是蛋白質與脂肪,尤其是以磷脂質(Phospholipid)為主之極性脂肪(Polarlipid)為主成分,大部分植物以不飽和脂肪酸為主。在種子含水量較低時,包為大分子的單層水膜成為不連續狀,而使氧及酵素與不飽和脂肪酸直接接觸如無抗氧化因子之保護,極易發生過氧化反應而產生自由基,自由基與鄰近的分子發生氧化還原作用使這些分子失去原來的功能,此可能是細胞膜被破壞分解使發生滲漏及降低細胞合成能力之主因。其反應如下:
- 細胞膜:蛋白質、脂肪(磷脂質)不飽和脂肪酸 → 過氧化反應 → 自由基自由基十氧 →氫化過氧化物
- 氫化過氧化物→ 羰基
- 羰基十蛋白質→酵素鈍化、膜的損傷、組蛋白變性
- 羰基十脂質→破壞膜的結構
- 羰基十核酸→染色體畸形與突變
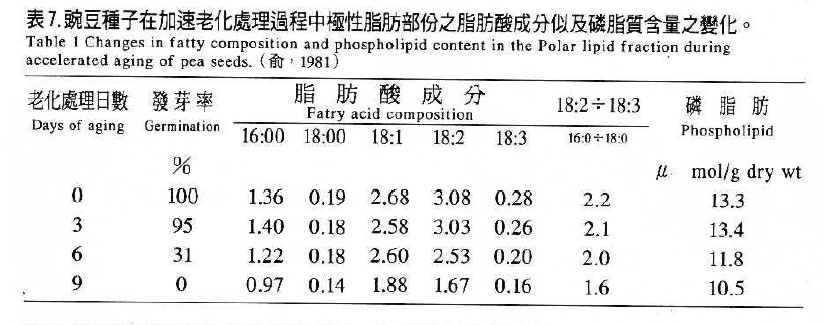
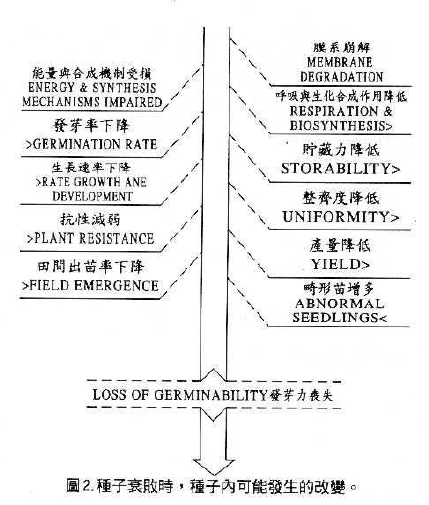
如表7(俞,1981)豌豆種子活力降低時,Linoleic acid (18:3)及Linoleic acid(18:2)含量先迅速下降,不飽和脂肪酸和飽和脂肪酸之比例亦迅速降低,磷脂肪之含量亦下降。在整個種子衰敗的現象大致可歸納成圖2(Delouche and Baskin,1973)。
(三)種子衰敗的特性(彭,1990)
- 種子衰敗是無可避免的。
- 種子衰敗是不可回覆的。
- 種子衰敗隨種子之種類;同種類不同批次;同批次中個別種子而互有差異。
(四)影響種子老化的因子:
1.水分:
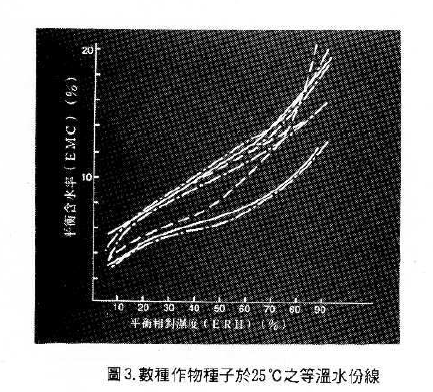
種子的水分含量與大氣中的相對濕度都會影饗種子老化,種子水分越高越容易老化,種子水分的有效區間是 5-7% ,因作物種類不同而異,例如花生為 2 %,豌豆為6.2 % ( Eillis , 1991)。相對濕度越高種子越易老化,一般相對濕度以不超過60%為宜。然而,種子是一種吸濕體(hydroscopic),它不斷的從大氣中吸收水分,又不斷的從自身中將水分蒸散到大氣中,直到達成平衛狀態,此時種子含水量稱為平衡含水量EMC (equilibrium moisture content),其相對濕度稱之為平衡相對濕度ERH。定溫下,EMC與ERH之關系曲線為等溫水分線(Isothems),如圖3在相同平衡相對濕度下,高澱粉含量的種子(高粱、小麥等),其含水量較高;含油量高的種子(大豆、萵苣)其含水量較低(陳與吳,1995)。一般增加種子貯藏壽命的平衡相對濕度在10%或水分潛勢(Watcer Potential )-350 MPa左右。
2.溫度:
高溫促進呼吸作用其生化活動產生的熱能足以傷害種子本身,而加速種子老化速率,不適的低溫對種子產生直接的傷害。溫度亦影響蟲害、黴菌的活動,溫度與水分含量有極密切的關係。一般而言,種子水分含量減少1%,溫度降低10℃下,貯藏能力增加兩倍。(溫度有效區間為50℉-60℉水分含量5-17 % , RH在 60%以下)(彭,1990)。
3.氧氣:
減少氧氣可以減少脂肪過氧化之機會,因此低氧環境可維持長久之種子活力,Siegal以抗氧化劑Cysteine處理菜豆,發現具有保護種子的功能(俞,1981 )。
四、促進種子活勢之處理技術:
(一)化學物質添加
1.殺菌劑:
種子表面常帶有複雜的微生物,其中有少許之病原菌在發芽過程中或種子田間萌芽致使種子腐敗或幼苗死亡。因此,添加殺菌劑或抗生素等處理後,可殺滅附於種子表面各種病原菌或孢子,對於種子壽命之延長或萌芽以及幼苗生長均有改善作用。例如以80 %得恩地(Thiram)可濕性粉劑或愛樂生粉劑(Arasan-50-red)於蔬菜種子(2-4g/kg)拌種處理。可增加豌豆、菜豆、蕪菁、萵苣、番茄幼苗對猝倒病之抗性(Jacks , 1951)。又例:十字花科種傳病害黑腐病可在播種前將種子浸泡在 52℃ 100µg/ml streptocy-cline ( 15% streptomycin+1.5% chlorot etracycline)溶液中 30 鐘可減少種子帶病源(Sharma,1980 )。
2 .養分:
以養分,微量元素甚至維生素添加處理,其中以缺乏某特定微量元素之處理最為有效,例如甜瓜經硼、鐵、鋅、錳、鋰或鉬鹽浸泡 24 小時及西瓜經鋅鹽溶液處理會促進發芽及生育;甜菜種子經適量的銅、錳及鋅鹽溶夜浸泡 24 小時後也會促進發芽(劉,1990)。
3 .荷爾蒙及植物生長調節劑:
荷爾蒙是控制種子活性機制的主要因子,胚活性減少與高濃度的 ABA、IAA、gibberellins 及 cytokinins 有關(朱,1981)。Gibberellins可以促進休眠及非休眠種子之發芽,因gibberellins 在種子發芽過程中扮演一極廣泛的角色(朱、王、陳,1981)。例如萵苣種子於播種前以cytokinins,及 GA 處理,可提升在高溫下之發芽率(Gray and Steckel,1976)。芹菜浸種於含有 GA4及 GA7 的混合物中與 Etheghon可改善發芽速率與發芽率(Thomas and Whitlock ,1980);南瓜與黃秋葵種子以 GA3處理會增加發芽率(Singh and singh,1973)。
(二)依生化觀點之種子處理技術
1 .種子造粒(披衣)( seed pelleting)處理:
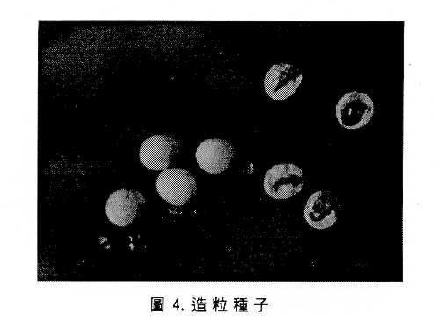
許多蔬菜種子體積小,形狀不規則,在播種時不易操作或機械化,因此在播種前即先行造粒處理,使用多種不同類型的矽藻土、親水性及疏水性的膠體為材料,使種子適於精密播種,通常在材料中可加入保護劑、營養劑或其它化學物質以改進發芽或幼曲萌芽能力(如圖4 )。
2.粉衣和膜衣處理( Coating and film coating ) :
粉衣和膜衣處理乃保持種子原來形狀和大小,加入殺蟲劑、殺菌劑、營養劑、生長調節劑,甚至於殺草劑等,於材料中可增加種子發芽率及萌芽後對環境的抵抗能力。而膜衣處理則在粉衣處理材料中加上聚合物和粘劑來處理種子(如圖5)。
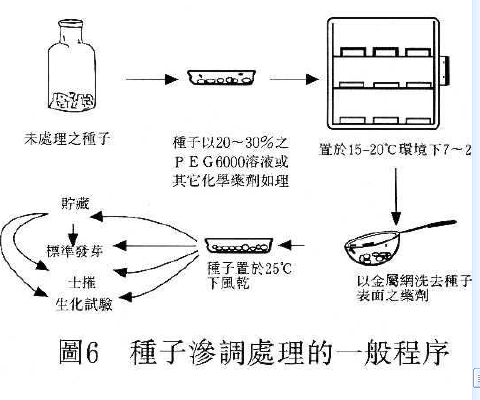
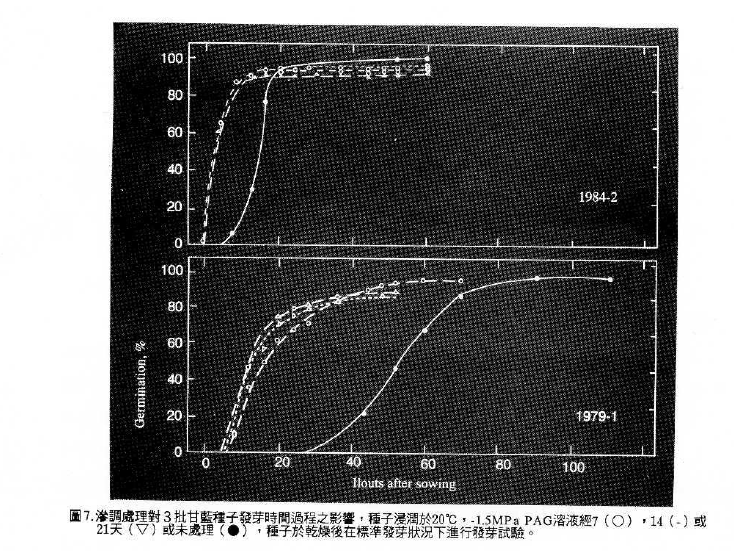
3.滲調處理( Priming , osmotic conditioning ) :
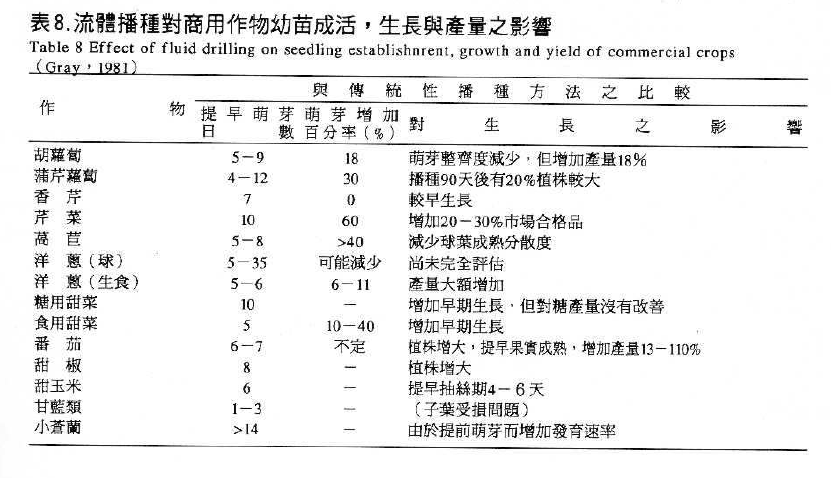
種子發芽過程中藉由浸潤溶液之滲透潛勢以延長種子吸水第二階段之停滯期,使種子能有充足的時問進行生理與生化之反應,並防止第三階段胚根突破種皮的發生,待除去試劑回乾後再行種植時,可縮短種子發芽過程在第二階段停滯期所準備的時間,得以獲得快且整齊的出曲現象或可延伸發芽溫度範圍,甚至促進老化種子活力的恢復。
相關照片
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 8 則留言
8 則留言
