

 6223
6223
種子滲調處理於菠菜生產栽培之應用
種苗改良繁殖場 黃玉梅、王小華、陳國雄
前言
菠菜為營養價值極高的重要葉用蔬菜之一,一般農民都以種子直播的方式生產。農民直播或商品上所稱的菠菜種子在植物學上屬於果實,依種子外所包覆的果皮形態不同而分為圓粒種及角粒種。其果皮組織由排列緊密的柵欄細胞(palisade cell)構成同時具有厚壁組織,造成機械障礙。且種子吸水後會產生膠質阻礙氣體通透進而影響發芽,種子浸潤時也容易有酚類化合物產生而抑制種子發芽(Atherton and Farooque,1983)。此外菠菜種子發芽亦明顯受溫度影響,發芽率隨溫度升高而降低(王等,1999),超過25℃以上發芽率即顯著下降,10℃以下則發芽日數長且有發芽不整齊的現象(沈,1995)。
因此,菠菜種子於田間播種時極易受土壤環境及氣溫影響,只要有輕微的逆境,就會影響種子的發芽率及萌芽之整齊性。雖然以人工剝除果皮或以硫酸、過氧化氫、激勃素(GA3)等化學方法處理均有提高范菜種子發芽率及發芽速度的效果(宋,1990),但應用於大量處狂及實際生產時,處理方法除必須具有方便性、穩定性及一致性外,對於短期葉菜類更須考慮用藥之安全性。因此,利用滲調處理來促進萌芽是一個較理想的方法。聚乙二醇(Polyethylene glycol.PEG)為化學惰性,無毒、穩定且分子量大之試劑(郭與朱,1981;Mexal et al.,1975),因此PEG滲調處理就成為最適的選擇。本試驗以不同滲透潛勢之PEG溶液,不同溫度以及日數之滲調處理。篩檢能增進菠菜種子發芽且能改善高溫逆境下之發芽率的最佳處理條件,以大量處理的方式改進菠菜之田間生產。
材料及方法
試驗材料:菠菜種子,參試品種有西螺一號、都拉益、菠麗得等三種。浸潤基質為聚乙二醇(polyethylene glycol PEG:分子量6000)。
試驗方法:
將PEG 6000依Michel & Kaufman(1973)方法配製成-0.8MPa、-1.1 MPa、-1.4 MPa、-1.7 MPa等不同滲透潛勢之滲調溶液,然後,以種子:滲調液為1:20之比例於10℃、20℃及30℃的生長箱內分別處理4、6、8、10、12天,並以打氣幫浦進行通氣。經滲調處理之種子以牴上法採三重覆,分別於15℃及30℃發芽箱內進行發芽試驗,播種後每日紀錄發芽數至第7日止,最後計算發芽率及達到最終發芽率之50%所需天數(GT50)。另將'都拉益'范菜種子以-0.8MPa之PEG滲調液在10℃生長箱內滲漏處理4天後,於15℃下進行回乾兩天,回乾種子於1999年八月十九日在西螺蔬菜產區以繖播方試進行田間試驗,試驗面積共0.1公頃,播種量為1.8公斤/0.1公頃。田區採逢機區集設計四重複,小區面積為8×10cm2,播種後45天後以坪割調查採收量。
試驗中滲調處理條件之處理溫度,處理日睥及滲調液之滲透潛勢等三個變因各組數據以三向複因子變方分析法(A three-way factorial anova)進行分析,其餘處理條件對發芽及滲調處理對產量等各組數據則經Duncan’s多變堿測驗測定其差異顯著性。
結果
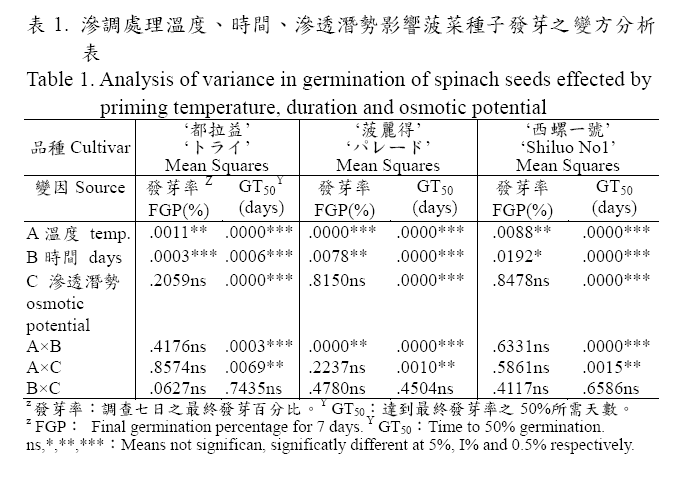
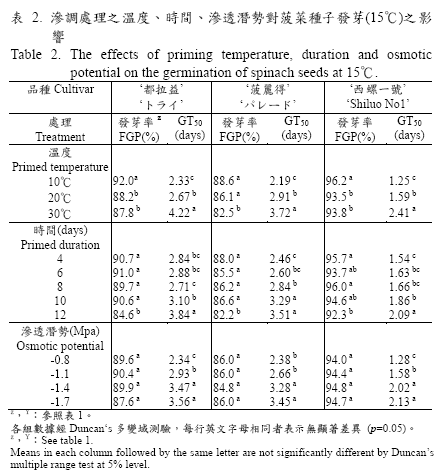
試驗之三個滲調處理條件中溫度、日數對菠菜種子在15℃下7天後之發芽率及達到最終發芽率之50%所需天數(GT50)均呈顯著差異,而不同滲透潛勢的PEG 滲調液處理對發芽率之影響則差異不顯著,但對GT50 則有顯著性影響(表1)。另外,滲調處理溫度與時間以及溫度與滲調液之滲透潛勢之間對GT50 形成顯著交感,也就是處理溫度對GT50 的影響會隨著處理天數和滲調液的滲透潛勢的變化而改變,且供試三個品種均有相同趨勢。從表2 得知:三個供試菠菜種子在不同滲調處理溫度(10℃,20℃,30℃)中,均以10℃之處理條件所得發芽率為最高,在88.6%-96.2%之間(因品種而異)且顯著高於其他兩個溫度,GT50 同樣以在10℃的處理條件為最短,在1.25-2.33 天之間(因品種而異),GT50 隨著處理溫度提高而增加,且不同處理溫度間呈顯著差異。
在處理時間方面:隨著處理天數增加,發芽率有逐漸降低的趨勢,五個處理天數中(4、6、8、10、12)以處理12 天之種子發芽率最低,而處理天數越少GT50 所需天數越短,其中又以處理4 天之1.54-2.71 天最短。另外,不同滲透潛勢的PEG 滲調液(-0.8MPa、-1.1MPa、-1.4MPa、-1.7Mpa)對發芽率的影響並不顯著,但對GT50則有顯著影響, PEG 滲調液滲透潛勢越高,種子發芽之GT50 越短,處理中以-0.8Mpa 滲調液得最快發芽(表2)。
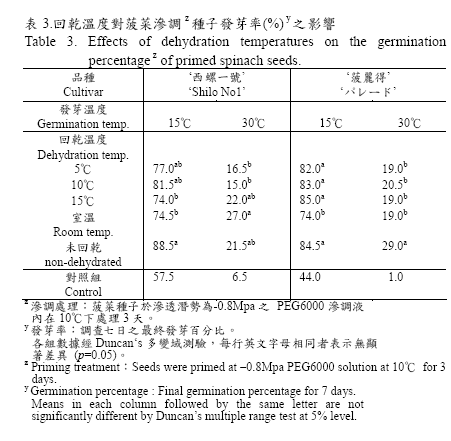
無論是否經過回乾處理程序,經滲調處理後的菠菜種子置於15℃及30℃之發芽溫度下,在調查七天後之發芽率均顯著高於未處理之種子(表3);唯供試兩品種在30℃的發芽環境下,經過滲調處理的種子發芽率雖較對照組的6.5%及1.0%顯著提高但均未能超過30%。另外,由表3 之試驗結果顯示:供試之‘西螺一號’種子於5℃、10℃下進行回乾處理與未回乾種子在適當的發芽溫度(15℃)下其發芽率顯著高於15℃及室溫下回乾的種子,但在高溫(30℃)發芽環境下,其結果則完全相反且較未回乾之種子為低。而‘菠麗得’種子於適當之發芽溫度(15℃)下之發芽率則以室溫回乾的74.0%為最低,其他於5、10、15℃進行回乾處理之種子與未回乾種子間之發芽率無顯著性差異,但在30℃高溫環境下所有經回乾處理之種子發芽率均顯著低於未回乾處理者。
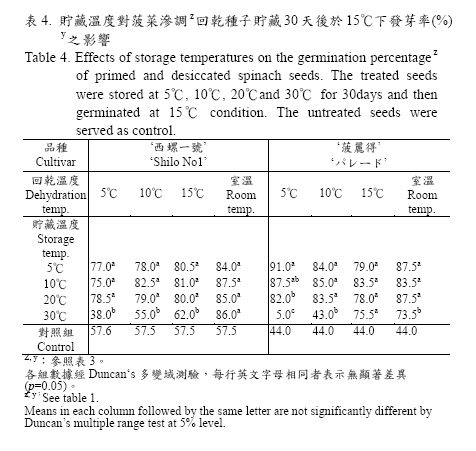
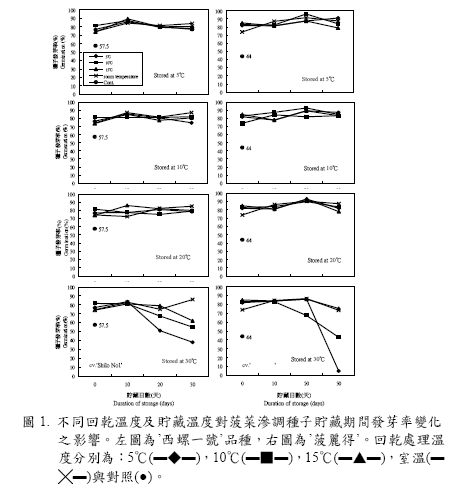
滲調後經不同溫度回乾處理之種子,以鋁鉑袋密封,分別於5、10、20、30℃環境下進行短期貯藏,由貯藏時間與種子發芽率之變化曲線(圖1)顯示:供試之‘西螺一號’及‘菠麗得’兩品種無論是在5、10、15℃或室溫下進行回乾的種子,在5、10、20℃環境下貯藏30 天後,其種子發芽率並無明顯下降趨勢,且顯著高於對照組的57.6%‘西螺一號’及44.0%‘菠麗得’。但是,貯藏於30℃環境下之‘西螺一號’種子(圖1 左下),在貯藏10天後種子發芽率即開始下降,其中又以低溫環境(5℃及10℃)回乾的種子降幅較大;貯藏30 天後只有室溫回乾的種子發芽率仍然維持不變,其餘5、10、15℃回乾的種子發芽率均已下降至70%以下(表4),且回乾處理溫度愈低,貯藏後的發芽率降幅愈大。另外,由圖1 右下之變化曲線顯示:在10℃回乾的‘菠麗得’種子於30℃貯藏20 天後發芽率已顯著下降,而5、15℃及室溫下回乾的種子發芽率尚能維持不變,但貯藏30 天後,於5℃及10℃回乾的種子發芽率則迅速下降,且比對照組的44.0%還低,其中又以5℃回乾之種子降幅較大,發芽率只有5%,而15℃及室溫下回乾的種子發芽率尚能維持在70%以上(表4)。
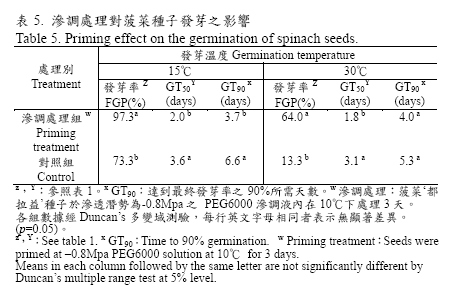
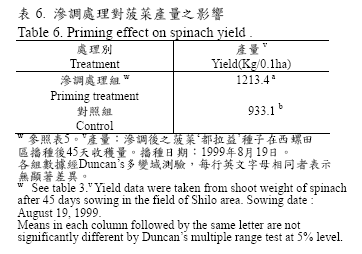
試驗中將菠菜種子以篩選之最佳滲調處理條件進行大量處理,處理回乾後之菠菜種子在15℃適溫下發芽率為97.3%,相同發芽條件下對照組之發芽率為73.3%,處理組GT50 為2.0 天而對照組則為3.6 天,GT90 處理組則較對照組提早3 天。在30℃高溫下經滲調處理之種子發芽率為64%對照組只有13%提高了51%,處理組GT50 為1.8 天而對照組為3.1 天,且兩者均呈顯著差異(表5)。該批種子同時於西螺蔬菜產區種植0.1 公頃,處理組播種後一週之萌芽情形即較對照組快速且整齊(圖2),於播種後45 天調查採收量,結果顯示經滲調處理者收穫量每0.1 公頃達1,213.4 公斤顯著高於對照組的933.1 公斤(表6)。
討論
滲調的主要目的在於加速種子萌芽、提高萌芽整齊度,尤其在不利於種子萌芽的環境中更加明顯(Khan et al., 1992;Parera and Cantliffe, 1994),此乃滲調處理後回乾之種子移入清水後,蛋白質和核甘酸合成加速,胚軸延長能力及時恢復,因此能迅速發芽(Bray et al.,1989),而種子滲調處理的成敗受到許多因素的影響,包括種子本身的品質及滲調處理期間通氣、光線、處理時間、溫度,滲調液的滲透潛勢以及滲透後的回乾與儲藏環境等(Parera and Cantliffe, 1994)。
通常採用PEG 液體滲調處理時,由於PEG 滲調液之溶氧較低,氧的移動性慢,必須靠外加通氣設備來增加溶液中的溶氧量,如洋蔥種子滲調處理之發芽率,外加通氣組即較不通氣組高(Bujalski et al., 1989),本試驗於滲調處理期間全程均以打氣幫浦進行通氣。另外,滲調時光線的外加與否,則視作物本身發芽期間對光的需求,如菊苣、萵苣、芹菜等好光性種子,於滲調期間照光處理,可提高種子發芽率或增加發芽整齊性(Geodes and Cantliffe, 1980;Bekendam et al., 1987),但菠菜非為好光性種子(劉,1988),滲調處理的溫度、時間與滲調液之滲透潛勢就成為影響菠菜種子滲調結果之重要變因。
適當的溫度有利於種子在發芽過程中吸水及進行生理、生化反應,使胚根能順利突破種皮,唯滲調處理的溫度往往低於作物的發芽適溫,如甘藍種子較適的滲調溫度為10℃,其發芽溫度則為20-30℃,其他如番茄、胡蘿蔔、洋蔥等,於15℃的滲調溫度下可得到較高的發芽率,而這些作物的發芽適溫在20-30℃之間(行政院農業委員會,1987;Khan, 1992;Khan et al., 1992),相同地菠菜種子發芽適當溫度為15℃(行政院農業委員會,1987),而本試驗三個滲調處理溫度中,也以較發芽適溫低的溫度條件(10℃),所得到的發芽率為最高且GT50所需天數最短(表2),此較發芽適溫低的滲調處理溫度條件,不但可有效的避免胚根突破種皮,且能延遲種子滲調時萌芽的時間,讓處理中的種子有足夠的時間,進行細胞膜修補工作及酵素的生合成(Powell and Mathews, 1981;Choudhuri and Basu, 1988)。另由表1 之變方分析顯示,三個處理變因對GT50 的影響中滲調處理溫度與處理時間及滲調液之滲透潛勢間的交感作用都是顯著的,因此為得到最佳之處理效果,除了滲調處理溫度外,必須同時考慮處理時間和滲調液的滲透潛勢的相互影響。
進行種子滲調時必須使種子在某限定的條件下(如低溫、低水分潛勢)浸潤吸水,使種子內原本近似靜止的各種生理代謝重新活化,但胚軸細胞的延展則受到浸潤基質(imbibitional medium)之低水分潛勢抑制,讓胚軸無法突破種(Heydecker and Gibbins, 1978),此時,滲調液的滲透潛勢及滲調處理時間成為調節種子浸潤之主因。而種子進行液體滲調處理時水分的吸收速度主要受到滲透潛勢的影響,而且滲調液的水分潛勢亦能影響滲調效果,當種子置於低的水分潛勢時,雖然種子內部與外界的水分潛勢達到平衡,但胚根的生長仍然受抑制(Bradford, 1986)。
試驗中菠菜種子在四個不同滲透潛勢(-0.8MPa、-1.1MPa、-1.4MPa、-1.7Mpa)的PEG 滲調液下處理,雖然其最終發芽率處理間並無顯著性差異,但在GT50 上則有顯著差異,其發芽所需天數隨著PEG 滲調液之滲透潛勢值提高而縮短,處理中以滲透潛勢-0.8MPa 發芽所需天數最少(表2),能得到最快速而整齊的發芽現象。另外,滲調處理所需時間乃必須讓種子吸水達到平衡,並足以活化種子內部的生理、生化反應,做好發芽前的準備工作,時間長短則因作物類別而異(Parera and Cantliffe, 1994),從表2 顯示本試驗供試三個菠菜品種之種子滲調處理時間不宜超過十天,否則會影響種子發芽率,而發芽GT50 以處理時間短的較少,其中又以處理四天之1.54-2.71天顯著少於其他處理天數。就試驗之因子間交感效應的分析結果顯示菠菜種子利用-0.8Mpa 滲調液在10℃溫度下處理四天所得之滲調種子發芽率最高且發芽速度最快。
種子滲調處理是否成功,除了包括種子水分的調控及滲調處理期間通氣、光線、處理時間、溫度,滲調液的滲透潛勢等因子,還必須含蓋滲調後的回乾與儲藏環境等(Parera and Cantliffe, 1994)。因此,回乾處理條件直接影響滲調處理之成敗及貯藏的時間。經滲調處理的種子必須使回乾到接近原來的水分含量,才能避免播種困擾並有利於短期貯藏,同時可以減少貯運時所遭受到物理性的傷害和微生物感染等問題(Khan and Ptasznik, 1992)。
經適當回乾處理之種子,仍然可以維持滲調之效果(Khan and Ptasznik, 1992),本試驗結果(表3)亦顯示經回乾處理後的菠菜種子在15℃及30℃之發芽溫度下,其種子發芽率均顯著高於未處理之對照組,證實回乾後的種子活力無論在適溫或高溫逆境下,仍可維持滲調之效果。又前人研究指出:回乾處理環境的溫度會影響回乾效益,而且所需條件因作物種類不同而有差異,其中適合較高回乾溫度者像是菜豆種子的35℃(Ptaszik and Khan, 1993)與萵苣種子的21℃;而適合較低回乾溫度者例如西瓜(Sachs, 1977)與洋蔥種子的15℃以及甘藍種子7℃(Perkins-Veazie et al., 1989)。
而針對菠菜種子的研究,學者僅以單一溫度進行回乾處理而未將不同回乾溫度加以比較,其中於常溫下乾燥回乾(Nakamura et al., 1982)及20℃回乾72 小時(Atherton and Farooque, 1983)等均不影響滲調效益,本試驗以四種回乾溫度(5、10、15℃、室溫)進行比較發現,回乾處理之溫度會影響種子之發芽勢表現,尤以室溫回乾之種子表現較差(表3),此現象是否與室溫回乾的處理條件使菠菜種子水分含量的下降速度較快有關亦或是其他原因,值得進一步探討。
前人在滲調種子回乾後之貯藏環境及貯藏時間與種子活力間的關係研究中,發現將經滲調回乾之菠菜種子於常溫乾燥條件下貯藏14 天後,其種子發芽率與未經貯藏之滲調種子間並無顯著性差異(Nakamura et al., 1982)。另將菠菜滲調回乾種子貯藏於5℃環境下30天後,其發芽率仍然可維持在87% (Atherton and Farooque, 1983)。本試驗將滲調後經不同回乾處理之菠菜種子置於5、10、20℃等三個貯藏環境30 天後,發現各種回乾處理之菠菜種子的發芽率在三個貯藏環境之間並無顯著性的差異表現,且發芽率均能維持滲調之效果(圖1),此與前人的研究結果有相同的趨勢。
但是在30℃的貯藏環境下,則種子的發芽率在四種不同回乾處理間呈現不一樣的變化,且品種間有些微的差異(圖1);原來僅回乾而尚未貯藏之種子發芽率是以較低溫度進行回乾處理之種子的表現較佳(表3),但是在高溫儲藏30 天後反而是以較高溫度進行回乾處理的種子表現最好(表4),也就是高溫的儲藏環境會使得種子的發芽率隨著回乾溫度的下降而降低且參試的兩個品種皆有相同趨勢,這種現象是否與回乾時的溫度與速度有關,為值得探究的有趣問題。
經滲調回乾處理之種子一旦再度吸水後,種子可快速而一致的發芽(Fu et al., 1988),試驗中滲調處理後回乾之種子,在15℃的適溫環境下較對照組提高24%,處理組發芽天數GT90 則較對照組提早3 天(表5),即明顯表現滲調之效果。滲調處理有助於高溫逆境下菠菜種子之發芽(Atherton and Farooque, 1983a;Atherton and Farooque,1983b),本試驗菠菜種子在30℃的高溫環境下經滲調處理之發芽率足足增加了51%,發芽天數亦明顯縮短(表5),利用PEG 滲調處理確實可以提高菠菜種子在高溫下的發芽率及加速種子發芽。
台灣中部地區菠菜產期依農民表示播種期最早在「白露」(國曆9月3 日),最晚至「清明」(國曆4 月5 日),平地在「清明」至「白露」期間,一般因高溫及早抽苔等問題而無法生產菠菜(王等,1998),農民多採用超量播種來彌補因高溫所造成的發芽率低落現象,中部地區在一般產期播種量為2.2Kg/0.1 公頃,高溫期將播種量增加到3.6Kg/0.1 公頃。本試驗採用‘都拉益’晚抽苔品種克服早抽苔現象,並利用篩選之最佳處理組合條件下進行大量滲調處理,使該批種子提早於「白露」前的8 月19 日在西螺蔬菜產區種植0.1 公頃,同時播種量減少為1.8Kg/0.1 公頃,而一般經滲調處理之種子出土率有被促進之效果(Alvarado and Bradford, 1988),其播種後一週滲調處理組之萌芽情形即較對照組快速且整齊(圖2),克服「白露」前高溫發芽困難的現象。試驗結果顯示菠菜‘都拉益’種子播種後45 天之採收量,經滲調處理者收穫量每0.1 公頃達1,213.4 公斤顯著高於對照組的933.1 公斤,其產量因滲調處理而提高。
綜上所述,菠菜滲調處理溫度以10℃所得種子發芽率最高且GT50 最短,滲調液濃度則以-0.8MPa 較佳。滲調之菠菜種子經回乾處理程序後,其種子發芽率仍可在短期內達到國家標準(CNS)的要求,且在貯藏30 天後發芽率仍可維持滲調的效果,唯受回乾處理與貯藏溫度的影響,處理條件中以15℃及室溫回乾及貯藏於5~20℃對維持菠菜滲調種子活力最為有利。本研究以種子發芽吸水曲線,配合滲調處理條件控制種子浸潤吸水速率,達到量產處理種子之目的,並將此技術實際運用於菠菜生產栽培上,改善菠菜種子萌芽整齊性及高溫逆境下發芽之限制,使在平地西螺地區能提早於八月播種,克服「白露」前高溫發芽困難的問題,產期提早後產值因而提高,直接增加農民收益。
參考文獻
1. 王三太、蕭吉雄、曹幸之、曾明懋。 1998。 雌雄異花同株菠菜品系之選育。 中華農業研究 47(1):54-62。
2. 王小華、鍾文全、沈再發。 1999。 溫度與滲調處理對菠菜種子發芽之影響。 植物種苗 1(1):1-6。
3. 中國國家標準。 1982。 蔬菜種子。 CNS 3317,N 1090 p.1-2 經濟部中央標準局印行。
4. 行政院農業委員會編譯。 1987。 國際種子檢查規則。 臺灣省政府農林廳印行p.140-167。
5. 宋 妤。 1990。 影響菠菜種子發芽因素之研究。 中國園藝36(4):280-289。
6. 沈再發。 1995。 菠菜 台灣農家要覽. 作物篇(二)。 豐年社發行p.331-334。
7. 郭華仁、朱 鈞。 1981。 種子滲調法。 科學農業 29:381-383。
8. 劉英德編著。 1988。 種子生理。 五洲出版社 p.243-283。
9. Alvarado, A. D. and K. J. Bradford. 1988. Priming and storage of tomato (Lycopersicum esculentum) seeds. I. Effects of storage temperature on germination rate and viability. Seed Sci. & Technol.16: 601-612.
10. Atherton, J. G. and A. M. Farooque. 1983. A. High temperature and germination in spinach. I. The role of the pericarp. Scient. Hort.19:25-32.
11. Atherton, J. G. and A. M. Farooque. 1983. B. High temperature and germination in spinach. II. Effects of osmotic priming. Scient. Hort.19:221-227.
12. Bekendam, J., J. G. van Pijlen, and H. L. Kkraak. 1987. The effect ofpriming on the rate and uniformity of germination of endive seed.Acta Hort. 215: 209-218.
13. Bradford, K. J. 1986. Manipulation of seed water relations via osmotic priming to improve germination under stress conditions.HortScience 21: 1105-1112.
14. Bray, C. M., P. A. Davison, M. Ashraf, and R. M. Taylor. 1989.Biochemical change during priming of leek seeds. Ann. Bot. 63:185-193.
15. Brocklehurst, P. A. and J. Dearmen. 1983. Interactions between seedpriming treatments and nine seed lots of carrot, celery and onion. Ⅱ.Seedling emergence and plant growth. Ann. Appl. Biol. 102: 585-593.
16. Bujalski, W., A. W. Nienow, and D. Gray. 1989. Establishing the large scale osmotic priming of onion seeds by using enriched air. Ann. Appl.Biol. 15: 171-176.
17. Choudhuri, N. and R. N. Basu. 1988. Maintenance of seed vigor and viability of onion (Allium cepa L.). Seed Sci. Technol. 16: 51-61.
18. Fu, J. R., X. H. Lu, R. Z. Chen, B. Z. Zhang, Z. S. Liu, Z. S. Li, and D.Y. Cai. 1988. Osmoconditioning of peanut (Arachis hypogea L.) seeds with PEG to improve vigor and some biological activities. Seed Sci.& Technol. 16: 197-212.
19. Geodes, A. M. and D. J. Cantliffe. 1980. Germination of lettuce seeds at high temperature after seed priming. J. Amer. Soc. Hort. Sci. 105:777-781.
20. Heydecker, W. and B. M. Gibbins. 1978. The priming of seeds. Acta Hort. 83: 213-223.
21. Khan, A. A. 1992. Preplant physiological seed conditioning. Hort. Rev.13: 131-181.
22. Khan, A. A., J. D. Maguire, G. S. Aabawi, and S. Ilyas. 1992.Matriconditioning of vegetable seeds to improve stand establishment in early field plantings. J. Amer. Soc. Hort. Sci. 117: 41-47.
23. Mexal, J., J. T. Fisher, J. Osteryoung, and P. C. P. Peid. 1975. Oxygen availability in polyethylene glycol and solutions its implications in plant-water relations. Plant Physiol. 55: 20-24.
24. Michel, B. E. and M. R. Kaufman. 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiology. 51: 914-916.
25. Nakamura, S., T. Teranishi, and M. Aoki. 1982.Promoting effect ofpolyethylene glycol on the germination of celery and spinach seeds. J.Japan. Soc. Hort. Sci. 50: 461-467.
26. Parera, C. A. and D. J. Cantliffe. 1994. Presowing seed priming. Hort.Rev. 16: 109-141.
27. Perkins-Veaznik, P. M., D. J. Cantliffe, and J. M. White. 1989.Improved stand establishment of direct-seeded cabbage with seed covers. J. Amer. Soc. Hort. Sci. 114: 36-39.
28. Powell, A. A. and S. Mathews. 1981. A Physiological explanation for solute leakage from dry pea embryos during imbibition. J. Exp. Bot.32: 1045-1050.
29. Ptaszik, W. and A. A. Khan. 1993. Retaining the benefits of matriconditioning by controlled drying of snap bean seeds.HortScience 19: 271-181.
30. Sachs, M. 1977. Priming of watermelon seeds for low-temperature germination. J. Amer. Soc. Hort. Sci. 102: 175-178.
相關照片
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 2 則留言
2 則留言
